- 移动端


武汉枢密脑科学技术有限公司品牌商
8 年
手机商铺
商家活跃:
产品热度:
- NaN
- 0.7000000000000002
- 1.7000000000000002
- 0.7000000000000002
- 3.7

抑制神经元活性Kir2.1:rAAV-hSyn-Kir2.1-EGFP
¥3980
品牌商
武汉枢密脑科学技术有限公司
入驻年限:8 年
- 联系人:
枢先生
- 所在地区:
湖北 武汉市 江夏区
- 业务范围:
抗体、试剂、细胞库 / 细胞培养、技术服务、耗材、实验室仪器 / 设备
- 经营模式:
生产厂商 经销商
推荐产品

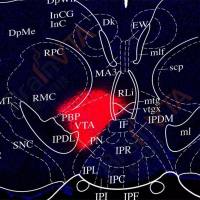

公司新闻/正文
神经元束路追踪技术
15987 人阅读发布时间:2023-08-04 11:14
大脑神经网络是由数目庞大,以及形态、特性各异的神经元,通过突触连接构成的复杂结构,是大脑行使认知、情感、记忆、想象等活动的结构基础。绘制神经元投射图能够追踪不同脑区之间的信息流动。下面我们主要讲解在实验动物中广泛使用的、用于确定不同脑区之间的连接方法。
一、 顺行示踪剂
顺行示踪剂主要被神经元胞体和树突吸收,然后运动至轴突,从而标记该神经元投射的区域。经典的顺行示踪剂主要包括以下几种类型:
1. 植物血凝素(PHA-L),这是一种在菜豆(Phaseolus vulgaris)中存在的凝集素。主要优点是所显示的神经纤维末梢形态非常细致,基本上没有过路纤维标记问题。
2. 放射性标记的氨基酸,这些氨基酸被吸收并掺入到神经元的蛋白质中,通过运动至轴突,也能够从轴突末端释放,并被对应的突触后神经元吸收。
顺行示踪剂有可能通过受体选择性被吸收到胞体,或通过非选择性的方式吸收,但由于胞体体积比轴突末端大得多,被富集到胞体。正向追踪通常与原位杂交、免疫染色或尼氏染色法一起使用。
二、 顺行示踪病毒
经典顺行示踪剂的局限在于,注射位点的所有细胞都会吸收示踪剂,因此这种方法所显示的投射模式是不同类型神经元投射的整体反映。大量的数据表明,同一区域不同类型的神经元具有不同的投射模式。另外,投射到同一靶区域的神经元也可能在该区域内具有不同的连接模式。对不仅在同一区域,而且属于特定类型的神经元的追踪将为绘制神经连接图提供更高的分辨率。
因此近年病毒作为神经元追踪试剂,既能在方向上做到更严谨,并能结合 Cre-LoxP 技术实现细胞类型特异性标记,这是病毒用作追踪剂所独具的特点。如在某些特定类型的细胞中表达了 Cre 重组酶的转基因小鼠中,通过注射包含依赖 Cre 表达的标记物的 AAV,可以实现特异细胞类型的正向追踪。
此外,这些病毒还具备携带基因工具(如光遗传工具、钙敏感染料、基因编辑工具、RNA 干扰工具等)的能力,其将在神经环路功能的研究中发挥重要作用。
常用的一些不跨突触的辅助病毒载体包括血清型为 2 型,8 型, 9 型的腺相关病毒、逆转录病毒和用 VSV-G 包装的慢病毒等,如下表: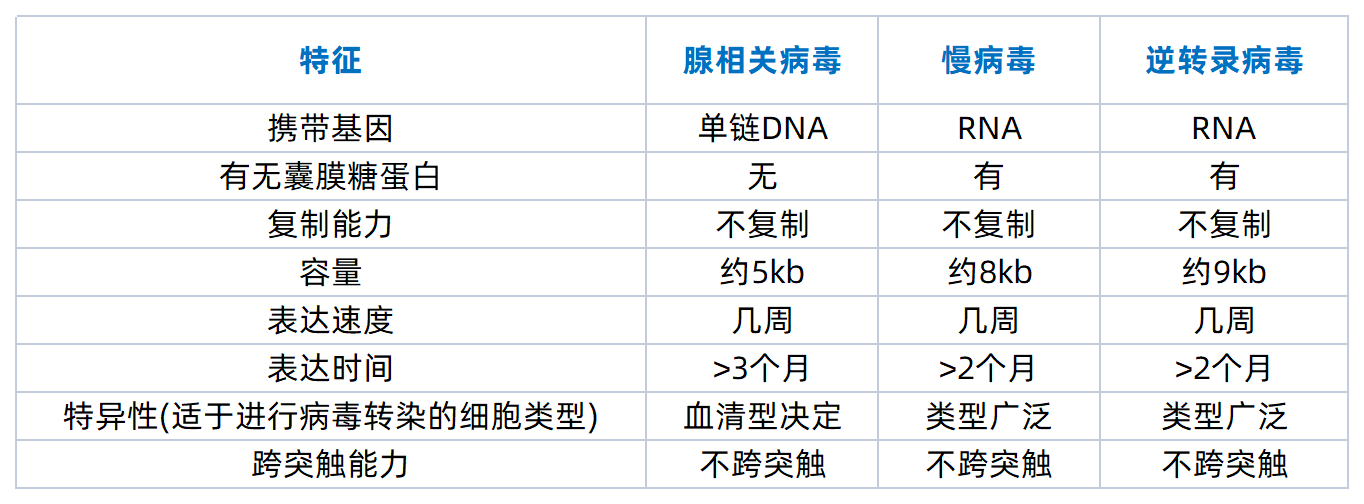
三、 案例展示
实例一 POMC 神经元全脑传出的鉴定
实验动物:POMC-Cre 小鼠
使用的病毒:AAV9-DIO-mtdTomato,AAV9-DIO-EmGFP
注射部位及病毒量:ARC 和 NTS,体积各 300 nl,滴度 2.00E+12vg/mL
检测方法:为较好的标记轴突,一个月后灌流取材,荧光纤维镜成像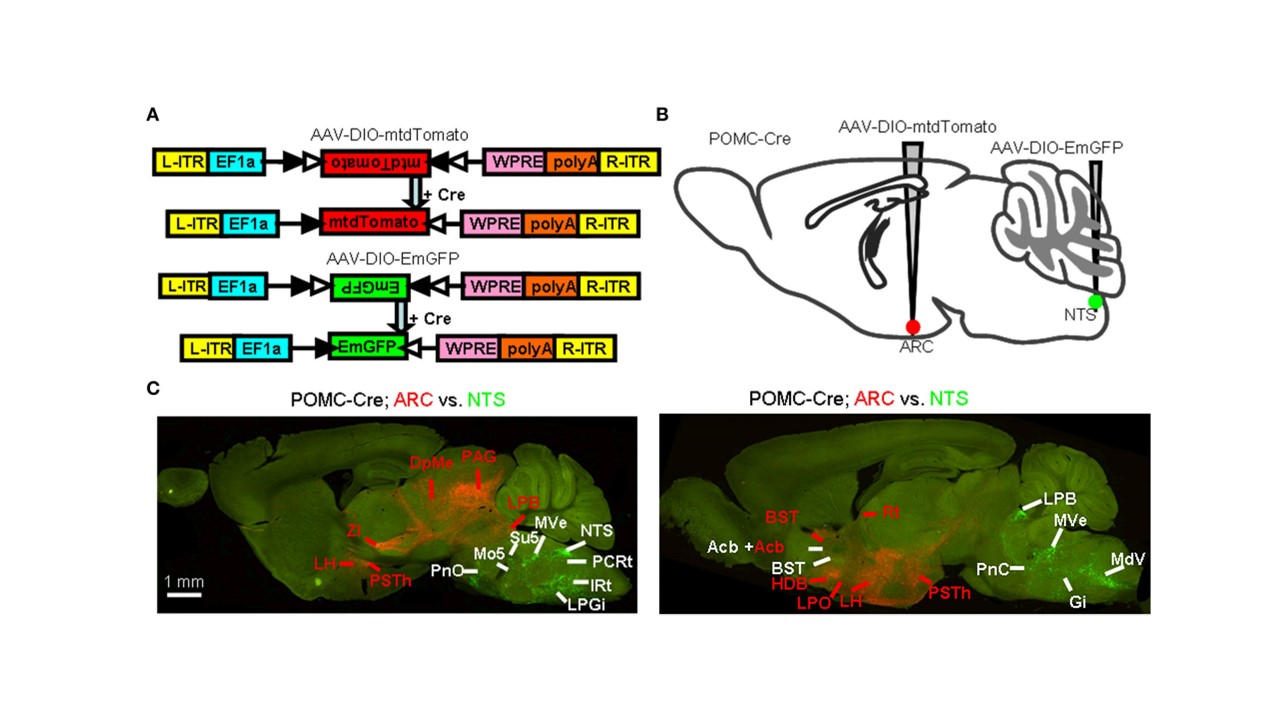
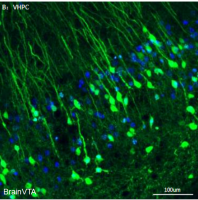
实例二 Dyrk1a(双特异性酪氨酸-(Y)-磷酸化调节激酶 1A)在脑发育中的作用
实验动物:出生前和出生后的小鼠
使用的质粒:慢病毒质粒
实验方法:宫内电穿孔注入 E14.5 小鼠脑室,P0 处死小鼠,取材,成像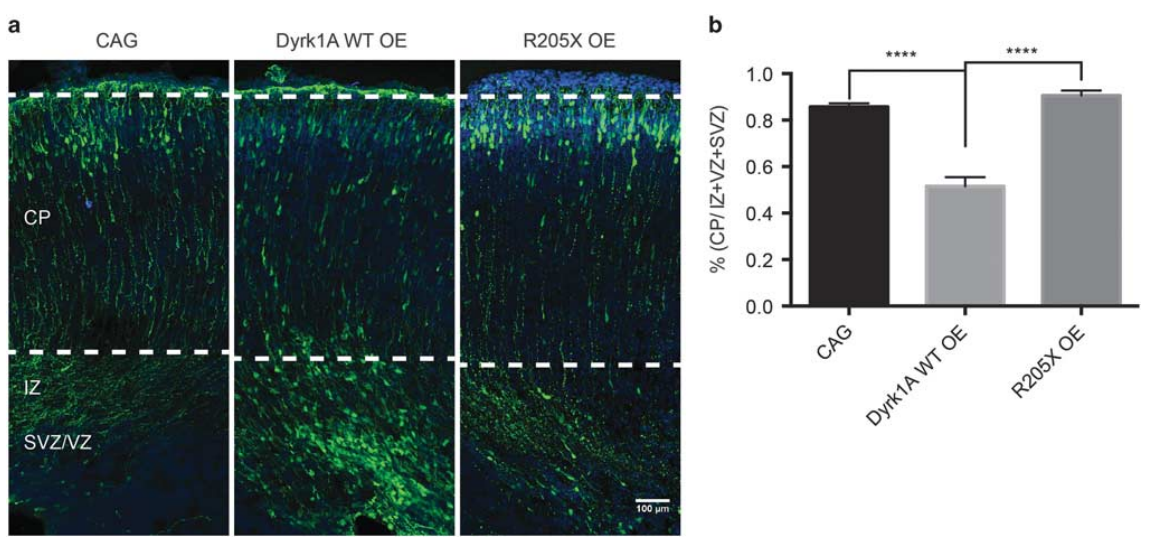
一、 逆行示踪剂
逆行示踪剂通常被轴突末端吸收然后反向运输到胞体。经典的逆行示踪剂包括:
1. 辣根过氧化物酶(HRP):1971 年 Kristenson 等及 1972 年 Lavail 等先后将 HRP(从辣根中提取的一组同功酶的混合物)用于追踪周围神经及中枢神经系统的纤维联系,此法的问世大大推动了神经解剖学的发展。它是基于神经元轴浆运输的生理现象,即将 HRP 注射至中枢神经系统或周围神经的一定部位,HRP 可被神经末梢以胞饮的方式摄入,逆行运送至胞体,经过一定时间后取相应的部位固定、切片,然后用组织化学方法显示 HRP 的标记。
2. 霍乱毒素亚基 b(CTb):霍乱毒素 (cholera toxin, CT) 是一种很灵敏的顺、逆向追踪剂。霍乱毒素有 A、B 两个亚单位,A 亚单位为毒素的毒性单位,B 亚单位为与细胞受体结合单位,无毒性,用 B 亚单位作追踪剂效果更佳。单独使用时的标记结果用抗 CTb 抗体的免疫组织化学方法显示。通常将其与 HRP 交联,形成 CT-HRP 或 CB-HRP,能很好地显示神经元的树突,直至其末端分支,大大提高了 HRP 追踪剂的灵敏度。
确定一个示踪剂究竟是正向还是逆向运输主要靠观察。逆行示踪剂通常与选择性富集在轴突末端的受体结合,通过轴突末端的内吞作用被吸收,通过内源的逆向轴突运输系统到达胞体。
二、 逆行示踪病毒
和经典的顺行示踪剂一样,上述传统的逆行示踪剂也具有方向不特异,不能实现同一靶区域内特异类型神经元的追踪。
常用的一些不跨突触的逆行病毒载体有:
1. 血清型为 Retro 的腺相关病毒 AAV。在 D.Gowanlock R.Rervo 的工作之前,经典的血清型对于神经元投射的逆向标记存在各种缺陷,不能满足各种科学研究的需求。rAAV/Retro 有广阔的应用前景。首先是逆行标记效率高:较经典血清型提高了 100 倍以上,甚至可以达到染料和 Retrobead 类似的逆标效率。其次是应用广泛:1)rAAV/Retro 可以轻松表达出所携带的目的基因,做以往很难实现的各种操纵;2)临床的应运中也有巨大潜力, 可以与其他的神经示踪病毒结合使用,特异、有目的解析神经环路问题,针对性地进行神经元群的基因编辑等多样性的基因治疗。
2. 缺失 RV-G 基因的重组狂犬病毒。Wickersham 等基于 RV 疫苗株 Sad B-19 感染性克隆构建的重组狂犬病毒,其 G 蛋白基因被敲除,并携带 GFP、mCherry 等荧光蛋白基因,可使外源蛋白在神经元中高丰度表达,从而清晰地标记神经元的精细形态。
3. 犬腺病毒2型(CAV-2)。CAV-2 病毒粒子主要侵染神经元的原因是由于柯萨基病毒和腺病毒的受体(CAR),该受体主要定位在神经元的突触前的位置。CAV 的神经元定位以及沿着轴突运输主要是 CAR 介导的。除了可以忽略的低免疫毒性以及对神经元的偏好性感染,CAV-2 病毒还是辅助病毒依赖的病毒粒子,拥有大约 30kb 的克隆装载量,这些基本性质扩展了 CAV-2 在中枢神经系统基因治疗中的作用。
三、 案例展示
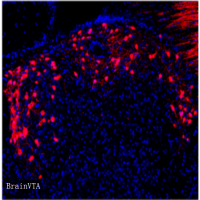
实例一 LC-SC 投射的逆行病毒示踪
实验动物:WT 小鼠
检测方法:AAV-Retro-Cre 注射在 SC,AAV9-DIO-mCherry 注射在 LC,4 周后可以在 LC 检测到 mCherry 荧光信号。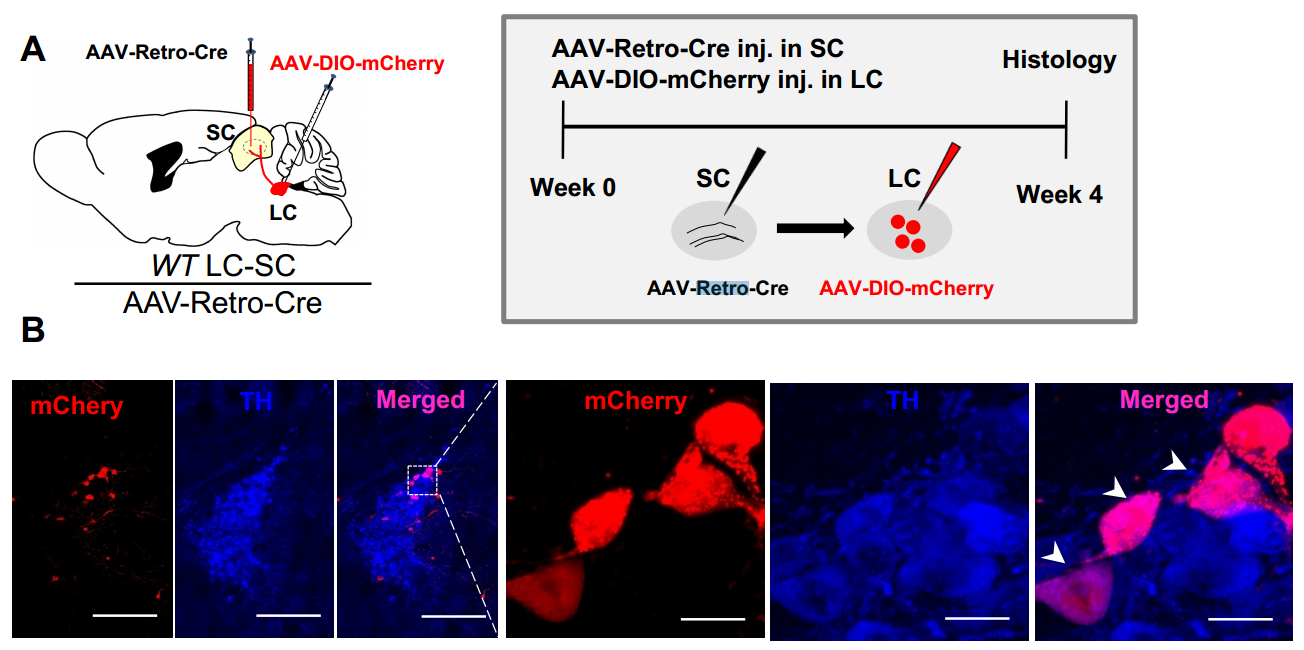
实例二 VTA 能神经元分离的支配 NAC 和 mOT
实验动物:C57BL/6 mice
使用的病毒:RV-DG-DsRed 和 RV-DG-GFP,滴度 10 的 8 次方 TU
实验方法和结果:分别注入 100 nl RV-DG-DsRed 和 RV-DG-GFP 在 NAC 和 mOT 中,1 周后灌流取材,可在 VTA 追踪到神经元。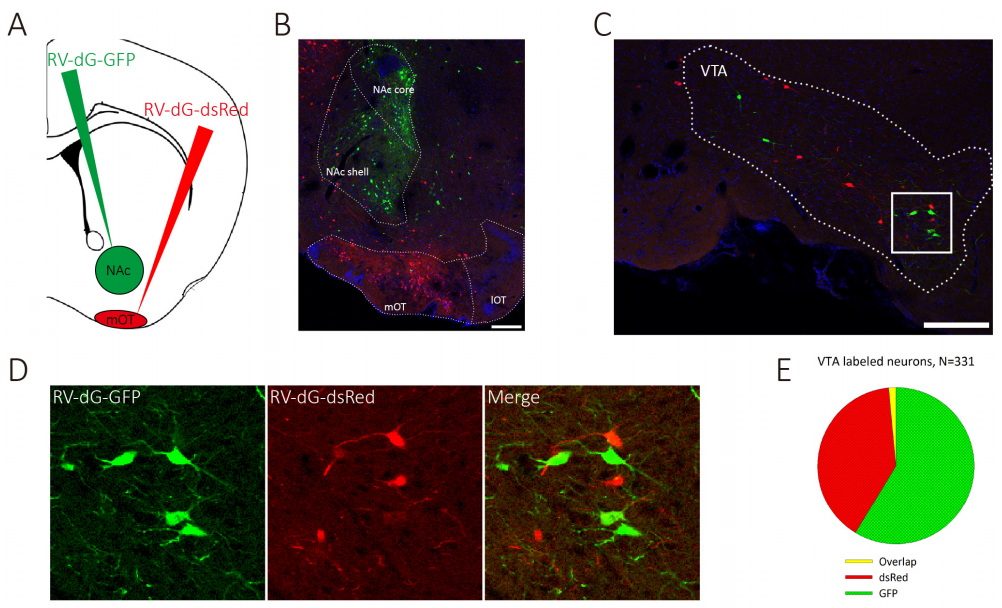
顺向追踪常用的方法
一、 顺行示踪剂
顺行示踪剂主要被神经元胞体和树突吸收,然后运动至轴突,从而标记该神经元投射的区域。经典的顺行示踪剂主要包括以下几种类型:
1. 植物血凝素(PHA-L),这是一种在菜豆(Phaseolus vulgaris)中存在的凝集素。主要优点是所显示的神经纤维末梢形态非常细致,基本上没有过路纤维标记问题。
2. 放射性标记的氨基酸,这些氨基酸被吸收并掺入到神经元的蛋白质中,通过运动至轴突,也能够从轴突末端释放,并被对应的突触后神经元吸收。
顺行示踪剂有可能通过受体选择性被吸收到胞体,或通过非选择性的方式吸收,但由于胞体体积比轴突末端大得多,被富集到胞体。正向追踪通常与原位杂交、免疫染色或尼氏染色法一起使用。
二、 顺行示踪病毒
经典顺行示踪剂的局限在于,注射位点的所有细胞都会吸收示踪剂,因此这种方法所显示的投射模式是不同类型神经元投射的整体反映。大量的数据表明,同一区域不同类型的神经元具有不同的投射模式。另外,投射到同一靶区域的神经元也可能在该区域内具有不同的连接模式。对不仅在同一区域,而且属于特定类型的神经元的追踪将为绘制神经连接图提供更高的分辨率。
因此近年病毒作为神经元追踪试剂,既能在方向上做到更严谨,并能结合 Cre-LoxP 技术实现细胞类型特异性标记,这是病毒用作追踪剂所独具的特点。如在某些特定类型的细胞中表达了 Cre 重组酶的转基因小鼠中,通过注射包含依赖 Cre 表达的标记物的 AAV,可以实现特异细胞类型的正向追踪。
此外,这些病毒还具备携带基因工具(如光遗传工具、钙敏感染料、基因编辑工具、RNA 干扰工具等)的能力,其将在神经环路功能的研究中发挥重要作用。
常用的一些不跨突触的辅助病毒载体包括血清型为 2 型,8 型, 9 型的腺相关病毒、逆转录病毒和用 VSV-G 包装的慢病毒等,如下表:
三、 案例展示
实例一 POMC 神经元全脑传出的鉴定
实验动物:POMC-Cre 小鼠
使用的病毒:AAV9-DIO-mtdTomato,AAV9-DIO-EmGFP
注射部位及病毒量:ARC 和 NTS,体积各 300 nl,滴度 2.00E+12vg/mL
检测方法:为较好的标记轴突,一个月后灌流取材,荧光纤维镜成像
图一. POMC 神经元全脑传出的鉴定。(Minmin Luo et al.Frontiers in Neuroanatomy.2015)
实例二 Dyrk1a(双特异性酪氨酸-(Y)-磷酸化调节激酶 1A)在脑发育中的作用
实验动物:出生前和出生后的小鼠
使用的质粒:慢病毒质粒
实验方法:宫内电穿孔注入 E14.5 小鼠脑室,P0 处死小鼠,取材,成像
图二 Dyrk1a (WT) 过表达可延迟小鼠胚胎皮质发育中神经元的迁移。(ZL Qiu et al. Molecular Psychiatry.2018)
逆向追踪常用方法
一、 逆行示踪剂
逆行示踪剂通常被轴突末端吸收然后反向运输到胞体。经典的逆行示踪剂包括:
1. 辣根过氧化物酶(HRP):1971 年 Kristenson 等及 1972 年 Lavail 等先后将 HRP(从辣根中提取的一组同功酶的混合物)用于追踪周围神经及中枢神经系统的纤维联系,此法的问世大大推动了神经解剖学的发展。它是基于神经元轴浆运输的生理现象,即将 HRP 注射至中枢神经系统或周围神经的一定部位,HRP 可被神经末梢以胞饮的方式摄入,逆行运送至胞体,经过一定时间后取相应的部位固定、切片,然后用组织化学方法显示 HRP 的标记。
2. 霍乱毒素亚基 b(CTb):霍乱毒素 (cholera toxin, CT) 是一种很灵敏的顺、逆向追踪剂。霍乱毒素有 A、B 两个亚单位,A 亚单位为毒素的毒性单位,B 亚单位为与细胞受体结合单位,无毒性,用 B 亚单位作追踪剂效果更佳。单独使用时的标记结果用抗 CTb 抗体的免疫组织化学方法显示。通常将其与 HRP 交联,形成 CT-HRP 或 CB-HRP,能很好地显示神经元的树突,直至其末端分支,大大提高了 HRP 追踪剂的灵敏度。
确定一个示踪剂究竟是正向还是逆向运输主要靠观察。逆行示踪剂通常与选择性富集在轴突末端的受体结合,通过轴突末端的内吞作用被吸收,通过内源的逆向轴突运输系统到达胞体。
二、 逆行示踪病毒
和经典的顺行示踪剂一样,上述传统的逆行示踪剂也具有方向不特异,不能实现同一靶区域内特异类型神经元的追踪。
常用的一些不跨突触的逆行病毒载体有:
1. 血清型为 Retro 的腺相关病毒 AAV。在 D.Gowanlock R.Rervo 的工作之前,经典的血清型对于神经元投射的逆向标记存在各种缺陷,不能满足各种科学研究的需求。rAAV/Retro 有广阔的应用前景。首先是逆行标记效率高:较经典血清型提高了 100 倍以上,甚至可以达到染料和 Retrobead 类似的逆标效率。其次是应用广泛:1)rAAV/Retro 可以轻松表达出所携带的目的基因,做以往很难实现的各种操纵;2)临床的应运中也有巨大潜力, 可以与其他的神经示踪病毒结合使用,特异、有目的解析神经环路问题,针对性地进行神经元群的基因编辑等多样性的基因治疗。
2. 缺失 RV-G 基因的重组狂犬病毒。Wickersham 等基于 RV 疫苗株 Sad B-19 感染性克隆构建的重组狂犬病毒,其 G 蛋白基因被敲除,并携带 GFP、mCherry 等荧光蛋白基因,可使外源蛋白在神经元中高丰度表达,从而清晰地标记神经元的精细形态。
3. 犬腺病毒2型(CAV-2)。CAV-2 病毒粒子主要侵染神经元的原因是由于柯萨基病毒和腺病毒的受体(CAR),该受体主要定位在神经元的突触前的位置。CAV 的神经元定位以及沿着轴突运输主要是 CAR 介导的。除了可以忽略的低免疫毒性以及对神经元的偏好性感染,CAV-2 病毒还是辅助病毒依赖的病毒粒子,拥有大约 30kb 的克隆装载量,这些基本性质扩展了 CAV-2 在中枢神经系统基因治疗中的作用。
三、 案例展示
实例一 LC-SC 投射的逆行病毒示踪
实验动物:WT 小鼠
检测方法:AAV-Retro-Cre 注射在 SC,AAV9-DIO-mCherry 注射在 LC,4 周后可以在 LC 检测到 mCherry 荧光信号。
图一. LC-SC 投射的逆行病毒示踪。(Liping Wang et al. Current Biology.2018)
实例二 VTA 能神经元分离的支配 NAC 和 mOT
实验动物:C57BL/6 mice
使用的病毒:RV-DG-DsRed 和 RV-DG-GFP,滴度 10 的 8 次方 TU
实验方法和结果:分别注入 100 nl RV-DG-DsRed 和 RV-DG-GFP 在 NAC 和 mOT 中,1 周后灌流取材,可在 VTA 追踪到神经元。
图二 TA 能神经元分离的支配 NAC 和 mOT。(Zhijian Zhang et al. eLife.2017)